
Characterizing Grizzly Bear Habitat using Vegetation Structure
In efforts to assess the population as well as potential management impacts, numerous studies have examined the role of anthropogenic features such as roads and seismic lines on grizzly bear home range size, movement patterns, and mortality. The impact of forest structure on populations of grizzly bears is less well understood, principally due to difficulties undertaking extensive field sampling of vegetation structure, or the use of poor surrogates of structure such as passive multispectral remote sensing datasets like landcover class or NDVI. These passive forms of remote sensing limits interpretation of vegetation structure due to its problems with spectral reflectances of different tree species, illumination angle effects, and shade. When continuous measurements are included in habitat selection studies, they are most commonly GIS-derived metrics such as distance to some feature of interest (e.g. road, forest edge) or DEM-derived topographic surface metrics, which do not directly measure forest structure. Lidar is an active remote sensing tool developed to provide highly detailed, three-dimensional representations of a scanned landscape through a combination laser-rangefinder and highly accurate, GPS-enabled inertial measurement unit (IMU). The arrangement of the “point cloud” has been shown to be related to the distribution of vegetation in the stand, allowing estimation of forest structure metrics.
In this study we investigated the role of vegetation structure in determining habitat selection at a fine-scale through the use of a matched case-control logistic regression, where sampling was conducted using a step-selection function (SSF).
Specifically we ask 2 questions. First, what is the role of canopy cover in interior forest stands on habitat selection? Second, do resources in interior forest stands modify the extreme selection for forest edges seen with grizzly bears? We hypothesized that the interaction between vegetation height and overstory cover could best explain habitat selection: we understand that habitat selection by bears is heavily influenced by forest structure, especially in the case of continental interior bears, and that we expect that when combined with information about watershed topographic moisture regime, solar radiation, and bears’ natural preference fore edge habitat (i.e. a core model), addition of height of dominant vegetation and overstory cover would improve model predictions.
Step Selection Function
We fit step an integrated selection function (iSSF) to model bear movement. The iSSF compares consecutive relocations (referred to as used steps) with some number of “control” relocations (also known as available steps) that are generated randomly from empirical or theoretical step length and turning angle distributions.
We defined a step as 3 consecutive hourly relocations to increase sample size, meaning our iSSF was simulating a correlated (steps generally proceed in a forward direction), unbiased (there’s no gravitation towards a specific feature) random walk. Turn angles were drawn from a Von Mises circular distribution (indicating generally forward movement) and control step lengths were drawn from a gamma distribution fit from empirically observed step lengths.
Variable Selection
We used Lidar to characterize forest structure using data acquired by the Government of Alberta during leaf-on conditions between 2007 and 2008. The point density was on average 1.2 returns/m2 and covered the eastern 21,000 km2 of the BMA. Data were processed using tools from FUSION and LAStools using an area-based approach according to lidar best practices. To best summarize forest structure using an area-based approach that avoids collinearity of predictors, metrics can be summarized in three general ways: measurements of height, measurements of coverage, and measurements of variability. Because forest age, canopy cover, and topographic indices (especially fine-scale topography) are good predictors for many food types; this was the foundation of our variable selection procedure. We used an iterative approach to variable selection where we first fit conditional logistic regression models for two classes of lidar variable (cover and height) to assess which metrics best captured broad scale selection of vegetation by bears. Considering the post-hoc nature of the analysis, we limited exploration of lidar variables to the following metrics:
| Cover | Percent all returns > 2.00 meters |
| Height |
95th percentile heights 75th percentile heights Mean height Quadratic mean height |
75th percentile of height was the top performing height variable, corresponding approximately to the mid-canopy (See figure 1).
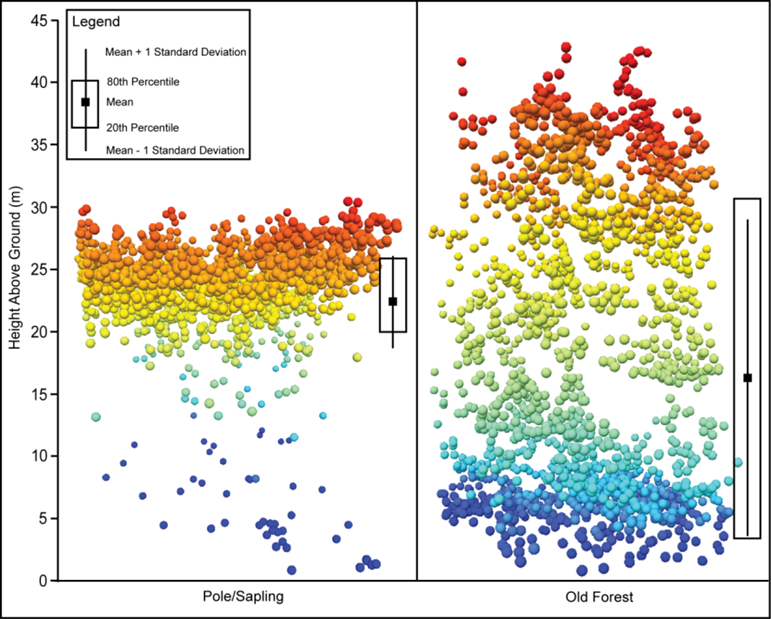
Figure 1 shows the way height percentiles can be interpreted
To quantify fine-scale topography we calculated the terrain wetness index (TWI, also known as compound topographic index), a measurement of topography that is highly correlated with soil moisture content, depth to soil horizons, and presence of vegetation.
Next, we included these variables in the context of other habitat descriptors. To minimize the chance of over-specification of the model, we limited contextual variables to those well linked to vegetation characteristics: elevation, solar radiation, and a measure of distance to forest edge. We defined forest using our lidar dataset, defined as areas with greater than 20% canopy cover and a dominant vegetative height (95th percentile height) greater than 3m. Because previous work in this area has shown that grizzly bears use roads with little regard for traffic volume and the vegetation structure of roadsides can be summarized with lidar height and cover metrics, we omitted roads from our model.
Model
Bears have strong seasonal patterns of feeding activity and the diversity of their diets is largely driven by vegetation phenology. Post-den emergence in the early part of the year, root digging and ungulate hunting/scavenging are dominant activities but they are abandoned in favor of frugivory once berries are present in quantity on the landscape. We used this knowledge to specify a priori that we would bin separate models corresponding to hypophagia (den emergence – 15 June), early hyperphagia (16 June -15 August), and late hyperphagia (16 August – den entry). Additionally, as bears are known to display different patterns of selection according to their sex, age, and reproductive status, we binned individual models according to sex-class (male, female, and females w/ cubs older than 1 year). We fit a conditional logistic regression to each individual’s GPS telemetry dataset and used AIC tally to select the best model (table 1 shows the breakdown of this division).
| Hypophagia | Early Hyperphagia | Later Hyperphagia | |
| Males | n = 7; 6 individuals | n = 10; 8 individuals | n = 11; 9 individuals |
| Females | n = 13; 9 individuals | n = 19; 10 individuals | n = 19; 11 individuals |
| Females w/ cubs | n = 4; 3 individuals | n = 4; 4 individuals | n = 3; 3 individuals |
| Table 1 Data breakdown, n indicates the number of “collar-years” of data available for analysis, the number of unique individuals is also indicated. |